par Prof. Dr. Paul Berth
Le blanchissement des coraux est un phénomène dont on entend souvent parler dans les médias. Il s’agirait d’un grave problème environnemental, dont la fréquence augmente, et qui pourrait mener à la perte totale des récifs coralliens. Le réchauffement climatique global, qui serait causé par l’augmentation de la concentration atmosphérique en CO2 est, bien entendu, pointé du doigt. Cependant, le blanchissement des coraux n’est-il pas un phénomène très ancien? Est-il seulement causé par des variations de température? Quel recul avons-nous à ce sujet? Une récente publication de Nicholas Kamenos et Sebastian Hennige, deux chercheurs anglais des Universités de Glasgow et d’Édimbourg, apporte de nouveaux éléments[1].
Tout d’abord, rappelons que les coraux blanchissent lorsqu’ils expulsent leurs algues symbiotiques, organismes unicellulaires appelés zooxanthelles, et que ce phénomène fait suite à un stress de nature physique, chimique ou biologique. Pour les stress physiques il y a par exemple la température inadaptée[2] ou une lumière trop ou trop peu intense[3]. Au niveau chimique nous trouvons l’effet de certains xénobiotiques (par exemple des herbicides[4]), mais également un pH ou une salinité inadaptée. Quant aux stress biologiques il y a bien entendu les attaques virales et bactériennes[5].
Mais ce que les alarmistes ne disent pas c’est qu’une fois devenus blancs les coraux ne sont pas encore morts… Le blanchissement serait en fait un moyen pour une colonie de polypes de s’adapter rapidement à un environnement fluctuant[6]. Les biologistes qui ont compris cela ont appelé ce phénomène « le blanchissement adaptatif ». Après l’épisode de blanchissement, les coraux acquièrent de nouvelles zooxanthelles, mieux adaptées au nouvel environnement. Il est vrai que la plupart des polypes du corail meurent, mais ceux qui survivent sont désormais plus résistants. En quelques mois, voire quelques années après le blanchiment, le corail a complètement récupéré car les polypes résistants se sont multipliés. Comme mentionné dans un article précédent, la théorie du blanchissement adaptatif a été très récemment confirmée par une étude génétique réalisée en 2016.
Mais quel recul avons-nous au sujet du blanchissement des coraux? Il faut savoir qu’avant 1979, très peu de choses étaient connues au sujet de la fréquence de blanchissement. Ceci est facile à comprendre : pour pouvoir se déplacer sous l’eau et constater que les coraux blanchissent, il aura fallu attendre l’invention du scaphandre autonome et le développement de la plongée sous-marine libre de type SCUBA. Comme l’élaboration du prototype du premier détendeur moderne n’a été faite qu’en décembre 1942, lorsque le français Jacques-Yves Cousteau rencontra l’ingénieur Émile Gagnan, il n’est pas étonnant que notre recul concernant les épisodes de blanchissement des coraux n’est pas très grand.
Les deux chercheurs anglais mentionnés en début d’article ont donc essayé de voir si le phénomène de blanchissement pouvait être observé dans le passé. Pour cela ils ont réalisé des carottages dans des coraux massifs de type Porites spp (Figure 1) vivant au niveau de la Grande Barrière de corail en Australie.

Les coraux durs et massifs du genre Porites (Figure 1) sont trouvés dans les récifs du monde entier. Au moins 7 espèces dominantes de Porites ont été identifiées (P. lobata, P. solida, P. lutea, P. australiensis, P. mayeri, P. murrayensis, et P. anae) et il a été estimé que les plus vieux Porites ont au moins 700 ans[7]. En d’autres mots, ces coraux n’ont pas été éliminés par les nombreuses périodes du passé où la température ou la salinité ont varié. La vitesse de croissance de ces coraux peut être déterminée en examinant les anneaux de croissance annuels dans le squelette en employant des rayons X (Knutson et al. 1972)[8]. Il a ainsi été déterminé que ces coraux peuvent croître à une vitesse d’environ 10 mm par an. Les Porites sont donc des enregistreurs précis des environnements marins du passé, du moins pour la zone de surface des océans. Par exemple, la composition en isotopes de l’oxygène du squelette d’aragonite de ces coraux peut donner des informations sur la température de surface de l’eau de mer ainsi que sur sa composition isotopique, qui est ensuite utilisée pour reconstituer la température des océans du passé[9].
Pour quantifier la fréquence et la prévalence des évènements de blanchissement du passé en employant des carottes de Porites, Kamenos et Hennige ont analysé la vitesse d’extension linéaire de ce corail. En effet, lorsque le corail blanchit, son squelette carbonaté en garde la trace car le dépôt d’aragonite est ralentit. Mais il est important de se rappeler que de multiples facteurs peuvent influencer la vitesse d’extension linéaire, comme la baisse de salinité à l’embouchure d’un fleuve, la turbidité (tempêtes), les changements de concentration en nutriments, etc. Il est donc important de considérer plusieurs coraux de la même espèce à des endroits différents.
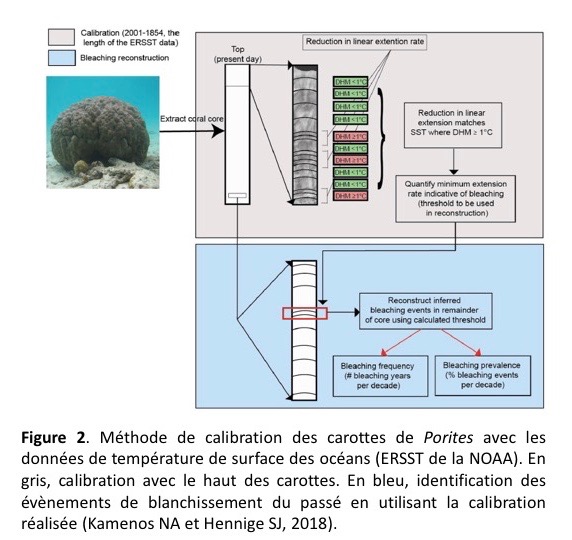
Les chercheurs ont analysé 44 carottes de corail et ont commencé par analyser le haut des carottes (la partie la plus récente). Pour cette partie haute, les stries de croissances ont d’abord été confrontées aux données de température de l’eau de mer (données ERSST fournies par la NOAA depuis 1854). Grâce à ces données de température de la NOAA, il est facile d’identifier les mois de l’année où la température locale moyenne était de 1°C supérieure à la moyenne habituelle. On sait que le blanchissement débute à ce moment. C’est ce qui est appelé une DHM ≥ 1°C (DHM, Degree Heating Month). Entre 1854 et 2001, c’est-à-dire la période couverte par les données ERSST, il y eut plusieurs DHM ≥ 1°C. Les chercheurs ont donc simplement mesuré la hauteur des stries du corail (i.e., la vitesse d’extension linéaire) au moment de ces épisodes DHM ≥ 1°C. Ils ont ainsi calibré leur outil.
Après cette calibration, les chercheurs ont regardé le bas des carottes de corail pour lesquelles aucune donnée de température ERSST n’était disponible, tout simplement parce que la NOAA n’existait pas avant 1854… Et ils virent que certaines stries de croissance avaient la même taille que celles se formant durant un épisode DHM ≥ 1°C. La procédure complète d’analyse est présentée en Figure 2.
En analysant les Porites massifs, Kamenos et Hennige sont ainsi remontés jusque 1620, un peu avant la Guerre de Trente Ans. C’est la première fois que l’on réussit à remonter si loin dans le temps. Bien entendu, pas de voitures et pas d’industrie pétrolière à cette époque, pas de pesticides et pas de surpêche, et donc si des épisodes de blanchissement sont détectés, ceux-ci doivent ipso facto être imputés à d’autres causes qu’à l’effet de l’homme. Rappelons qu’en 1620 les européens étaient à peine arrivés en Australie. Le Néerlandais Willem Jansz était le premier arrivé, en 1606. Son bateau, le Duyfken, jeta l’ancre devant cap York. Dans un récit postérieur, un Néerlandais décrit le territoire qu’il voit « comme non cultivé, et peuplé par de sauvages barbares noirs et cruels, qui ont massacré certains de nos marins »[10]. Les premiers explorateurs britanniques, William Dampier et James Cook, n’arrivent que plus tard entre 1688 et 1770.
Voici maintenant les résultats de Kamenos et Hennige : ils démontrent clairement que le blanchissement n’est pas un phénomène moderne. Les coraux blanchissaient en 1620 et la fréquence de blanchissement (le nombre d’années de blanchissement par décade) a même augmenté entre 1620 et 1753, pour ensuite diminuer de 1754 à 1820. A partie de 1820 jusque 2001, la fréquence augmente à nouveau, et ce à une vitesse comparable à la période croissante précédente (Figure 3).

Conclusions
- Le blanchissement des coraux existait avant l’ère industrielle et est apparu déjà en 1620.
- La fréquence de blanchissement peut augmenter ou diminuer en fonction du temps.
- L’augmentation actuelle de la fréquence de blanchissement n’est pas exceptionnelle.
- L’effet de l’homme sur la fréquence de blanchissement semble limité, du moins pour les coraux du genre Porites.
Références
[1] Kamenos NA and Hennige SJ (2018) Reconstructing Four Centuries of Temperature-Induced Coral Bleaching on the Great Barrier Reef. Front. Mar. Sci. 5:283. doi: 10.3389/fmars.2018.00283.
[2] Saxby T, Dennison WC, Hoegh-Guldberg O (2003). « Photosynthetic responses of the coral Montipora digitata to cold temperature stress ». Marine Ecology Progress Series. 248: 85–97.
[3] Hoegh-Guldberg O, Smith G (1989). « The effect of sudden changes in temperature, light and salinity on the population density and export of zooxanthellae from the reef corals Stylophora pistillata Esper and Seriatopora hystrix Dana ». Journal of Experimental Marine Biology and Ecology. 129 (3): 279–303.
[4] Jones RJ, Muller J, Haynes D, Schreiber U (2003). « Effects of herbicides diuron and atrazine on corals of the Great Barrier Reef, Australia ». Marine Ecology Progress Series. 251: 153–167.
[5] Kushmaro A, Rosenberg E, Fine M, Loya Y (1997). « Bleaching of the coral Oculina patagonica by Vibrio AK-1 ». Marine Ecology Progress Series. 147: 159–65.
[6] Graham NA, Jennings S, MacNeil MA, Mouillot D, Wilson SK (February 2015). « Predicting climate-driven regime shifts versus rebound potential in coral reefs ». Nature. 518 (7537): 94–7.
[7] Potts, D.C.; Done, T.J.; Isdale, P.J.; Fisk, D.A. (1985). « Dominance of a Coral Community in the Genus Porites Scleractinia ». Marine Ecology Progress Series. 23 (1): 79–84.
[8] Knutson, D. W., Buddemeier, R. W., and Smith, S. V. (1972). Coral chronometers: seasonal growth bands in reef corals. Science 177, 270–272. doi: 10.1126/science.177.4045.270.
[9] Thompson, D. M. « Comparison of observed and simulated tropical climate trends using a forward model of coral δ18O ». Geophysical Research Letters. 38.
[10] Flannery, Tim; The Explorers; Text Publishing; Melbourne; 1998.